
Donax trunculus. Animal acuático marino de agua salada. Invertebrado, no tiene esqueleto interno. Suspensívoro, se alimenta de materia orgánica y microorganismos en suspensión en el agua. Bentónico, habita íntimamente relacionado con el fondo acuático. Infaunal, vive enterrado dentro del sedimento del fondo marino, salvo en la fase larvaria que durante unas pocas semanas vive sin enterrarse por encima del fondo, pelágico.
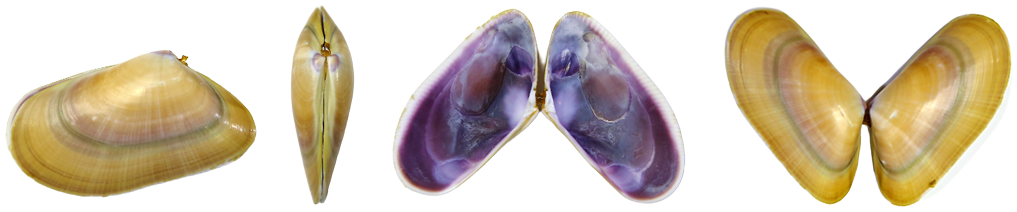
Imágenes 2.1. Diversas perspectivas de la concha, de las valvas, de un mismo individuo. Ejemplar de Valencia. España. Mar Mediterráneo. ©Jorge C. Tamayo Goya

Imágenes 2.2. Especie que en función de la distribución geográfica, de la variante, presenta diversos perfiles, no muy dispares entre sí, como por ejemplo los que aquí se muestran. Valvas de distintos individuos. Ejemplares de Valencia. España. Mar Mediterráneo. ©Jorge C. Tamayo Goya
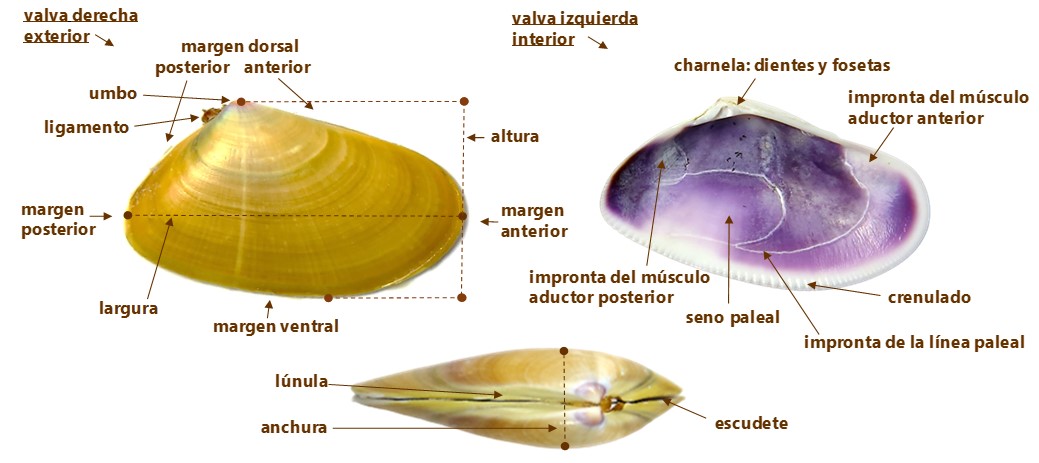
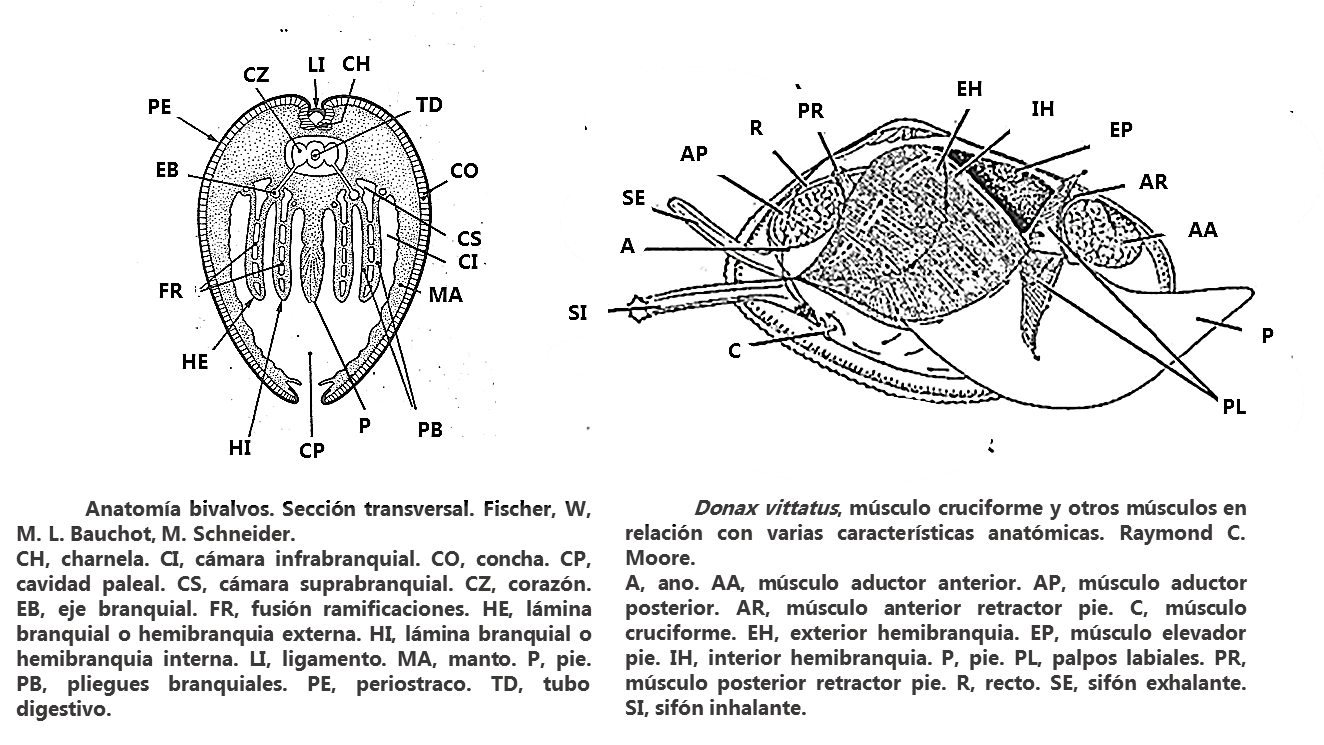
• La concha. Sólida, moderadamente convexa, cuneiforme. Inequilátera, ambos lados del plano umbo-ventral desiguales. Ligeramente inequivalva, valvas desiguales, la derecha es un poco más pequeña. (Imágenes 2.3) El borde dorsal anterior es un poco más alto en la valva izquierda, a la altura de la lúnula, que en el de la derecha y el margen dorsal de la valva izquierda sobrepasa ligeramente al de la derecha. Región anterior del margen dorsal, recta e inclinada. La región posterior más corta, oblicuamente truncada y algo convexa o más o menos rectilínea. Lúnula, es la pequeña depresión situada en la parte dorsal, anterior al umbo, alargada, profunda y lanceolada y escudete o corselete, es la depresión de la zona postero dorsal, inapreciable.
Margen anterior, redondeado. Margen posterior, más pequeño. Margen ventral, ligeramente convexo hacia el extremo anterior y casi paralelo al dorsal en la zona posterior Es posible que la unión de ambas valvas en el margen ventral anterior sea un poco flexuosa. Borde interno del margen ventral uniformemente crenulado, pequeñas muescas dentadas de poca profundidad, que se desvanecen en el margen anterior y desprovisto en el margen posterior o casi en su totalidad.
Superficie, brillante, lisa, aunque con algunas líneas concéntricas, estrías o pliegues, grietas o surcos, del crecimiento, algo más marcados en la región posterior. Líneas radiales, muy finas, más visibles al microscopio. Coloración exterior, variable, las tonalidades en marrón y en beige son predominantes, con frecuencia en tonos verde oliva, amarillento mostaza, también hay ejemplares en tonos grisáceos y blanquecinos. Con bandas concéntricas de diversos colores y tonalidades, como el violeta. Con frecuencia con franjas radiales que parten del umbo y se ensanchan hacia la zona ventral, en diversas tonalidades haciendo contraste. Interior, blanco, con áreas más o menos extendidas de color violeta o anaranjado, menos frecuente este último color que se desarrolla a partir del umbo, en cuyo caso es posible que también sea visible por el exterior pero en menor extensión.
La coloración se debe a distintos pigmentos que se fijan a la calcita en el ostraco, principalmente, y también está ligada a las condiciones medioambientales antes mencionadas. Estos pigmentos, melaninas y carotenoides, al ser de naturaleza orgánica, son sensibles a la luz, temperatura, humedad, etc., por lo que son alterables cuando el molusco se encuentra fuera de su hábitat.
Imágenes 2.3. Detalle del exterior e interior de valvas que no corresponden a la misma concha. Ejemplares de Valencia. España. Mar Mediterráneo. ©Jorge C. Tamayo Goya
Orientación de las valvas de Donax trunculus: /Apoye la parte exterior de una de las valvas sobre la palma de la mano con el margen anterior, el extremo más redondeado (por donde se extiende el pie), en dirección a la punta de los dedos y con el umbo, la protuberancia del dorsal superior, en dirección al pulgar. La orientación, la dirección de esa valva será la misma que la de esa mano. / La dirección del umbo, de la cavidad de la impresión del seno paleal y la ubicación del ligamento, indican el lado posterior. / Los sifones están ubicados en el lado posterior y el pie en el anterior.
(Imágenes 2.3) El umbo, parte sobresaliente o protuberancia dorsal, poco saliente, redondeado y opistogiro (dirigido hacia la parte posterior). Es en la que se inicia el crecimiento de la concha, el crecimiento de ambas valvas. En la región dorsal junto al umbo se encuentra la charnela, que tiene un interés relevante desde el punto de vista taxonómico, Heterodonta en el caso de Donax trunculus. (Imagen 2.4) La función de la charnela es evitar el movimiento o deslizamiento lateral. Está formada por prominencias (dientes) y huecos (fosetas), en el que los dientes de una valva encajan en las fosetas de la otra. La valva derecha con dos dientes cardinales, el posterior es bífido y el anterior simple, un diente posterior lateral y un diente lateral anterior. La valva izquierda con dos dientes cardinales, el anterior es bífido y el posterior solitario; un lateral anterior débil y un diente posterior lateral bien desarrollado.
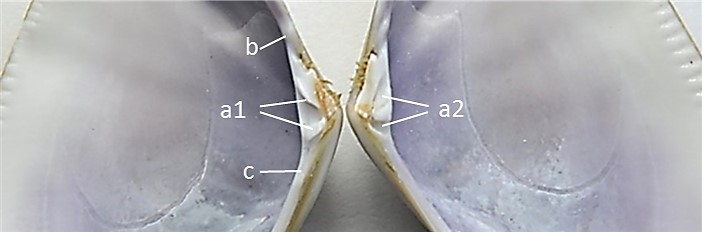
Imagen 2.4. Detalle de charnela, dientes y fosetas. Dientes cardinales, los dientes de una valva (a1) encajan en las fosetas (a2) de la otra. Diente posterior lateral (b). Diente anterior lateral (c). Ejemplar de Valencia. España. Mar Mediterráneo. ©Jorge C. Tamayo Goya
El ligamento (Imágenes 2.5), denominado tensilium cuando es externo y visible con las valvas cerradas, es una estructura cornea membranosa elástica marrón que une las dos valvas. Corto, situado en el borde dorsal posterior de la charnela, sobresale al exterior. Se aloja en la ninfa, área que refuerza la estructura de la charnela, una en cada valva. La concha tiende a mantenerse abierta por tracción del ligamento y distensión de los músculos aductores, los músculos más grandes y fuertes, desempeñando así una función antagónica a dichos músculos que la cierran cuando se contraen y el ligamento se relaja. Las impresiones o huellas que dejan en la concha son otro elemento característico de la especie.
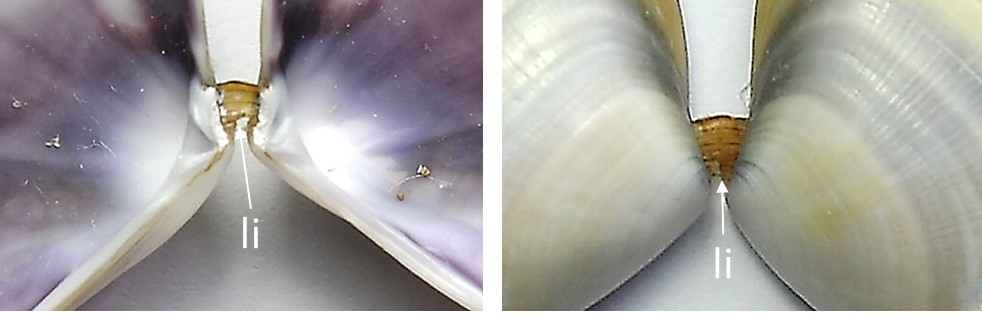
Imágenes 2.5. Ligamento (li). Ejemplar de Valencia. España. Mar Mediterráneo. ©Jorge C. Tamayo Goya
La concha es una estructura de una importancia adaptativa esencial para la defensa del animal que además sirve como anclaje y soporte de las partes blandas del cuerpo. Es el exoesqueleto o esqueleto externo del molusco. Está formada por dos partes articuladas, valvas, unidas merced al ligamento y a la charnela. Compuesta principalmente por carbonato de calcio que lo extraen del alimento que ingieren, es un depósito inorgánico que el pliegue interno del borde del manto transforma en cristales en forma de aragonito y calcita depositándolo formando capas. Está segregada por el epitelio del manto, que es una capa unicelular que reviste al manto.
La concha la integran, básicamentemente, tres capas superpuestas. (Imagen 2.6) Las dos principales, ostraco e hipostraco (Imágenes 2.7 y 2.8), están integradas a su vez por varias capas compuestas primordialmente por aragonito, calcita y en menor medida por conquiolina. La interior, hipostraco, miostraco o endostraco, capa en la que se fijan los músculos, formada por una estructura laminar cruzada, horizontal, espesa, proporcionando un aspecto porcelánico. La intermedia, ostraco o mesostraco, dispuesta de forma prismática constituye el principal soporte. La exterior, periostraco o ectostraco, (Imágenes 2.9 y 2.10) es la capa no calcárea que envuelve la concha, de naturaleza quitinosa, compuesta principalmente de conquiolina, parecida a una membrana, muy fina, muy adherente, translúcida, marrón verdosa, amarillenta, que protege las capas calcáreas de los ácidos corrosivos y de la fijación de otros organismos a la concha. Puede estar ausente por la acción erosiva del medio marino o cuando está muerto el molusco.
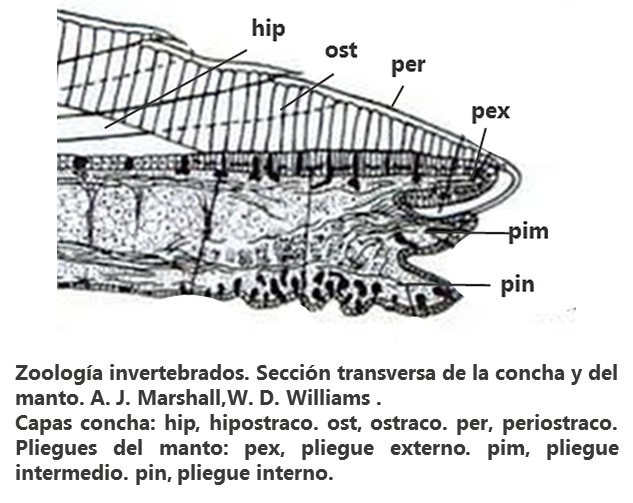
Imagen 2.6

Imágenes 2.7/A, 2.8/B, 2.9/C y 2.10/D. Detalle de las diversas capas que forman la concha: ostraco (os), hipostraco (hi) y periostraco (pe). Ejemplares de Valencia. España. Mar Mediterráneo. ©Jorge C. Tamayo Goya
Son capaces de reparar la concha si esta resulta dañada, siempre que la parte del manto que está debajo permanezca intacta, no deteriorada. Las conchas aragoníticas, como las de Donax trunculus, son resistentes a la fractura, pero cuando se produce se extiende irregularmente por la concha, y se puede apreciar mejor que se trata de un material cristalino.
La concha, las valvas, crecen tanto en superficie como en grosor. Crece más en longitud que en altura o anchura. Crece desde el dorso hacia el margen. Durante la ontogenia, período embrionario, el crecimiento en anchura es cada vez más importante que el crecimiento en longitud. El incremento superficial está en función del incremento de la superficie del manto, mientras que el incremento en grosor depende del índice de depósitos de carbonato cálcico y de matriz orgánica. La concha no crece de manera constante. Crece según las condiciones ambientales, temperatura, alimento, calidad del agua, etc. La concha es un registro medioambiental continuo.
En las especies de bivalvos se tiene la creencia de que varios son los factores ambientales que influyen en la morfología de la concha y las proporciones relativas, tales como, la profundidad, los niveles de la costa y de las mareas, la exposición a las olas, el tipo de fondo y tipo de sustrato. Se puede suponer que el crecimiento, desarrollo y supervivencia de los bivalvos están generalmente condicionados por parámetros físicos y químicos.
Las mayores tasas de crecimiento se dan en los ejemplares de las costas atlánticas de Galicia, aunque con un crecimiento más lento y un período reproductivo menos amplio que los del Mediterráneo. Los ejemplares de las costas gallegas del Atlántico son, de media, más grandes y con el extremo posterior más largo que las del Mediterráneo. Las aguas más cálidas favorecen el incremento de la cantidad de agua filtrada a través de las branquias, por lo que filtran más alimento, lo que se traduce en mayor capacidad de nutrirse. En el Mediterráneo, en el que la temperatura del agua rara vez cae por debajo de 12º, las poblaciones tienen un período reproductivo más amplio que las del Atlántico, en cuya zona intermareal están expuestas hasta 6º en invierno. Para su normal desarrollo las temperaturas del agua del mar u océano no deben estar por debajo de 5º y no alcanzar los 30º.
Durante los primeros dos meses y medio de vida experimentan un rápido crecimiento y en cinco o seis meses pueden alcanzar la mitad de su tamaño total. Tanto en el Mediterráneo como en el Atlántico, al final del primer año de vida muchos individuos pueden alcanzar 25 mm de longitud, entre 32-36 mm al final del segundo-tercer año y entre 37-42 mm en el tercer o cuarto año, dependiendo de la zona geográfica. Las mayores tasas de crecimiento se dan en las costas atlánticas de Galicia, como se ha señalado antes. En verano es cuando experimentan mayor crecimiento, mientras que en otoño-invierno se produce una desaceleración con lento crecimiento, e inclusive hasta puede casi detenerse en algún momento.
Tallas más comunes en el Mediterráneo, entre 25 y 35 mm., excepcionalmente por encima de 50 mm; en el Atlántico, entre 30 y 42 mm., rara vez por encima de 52 mm.

Imagen 2.11. Individuo de 44 mm de longitud. Ejemplar de Valencia. España. Mar Mediterráneo. ©Jorge C. Tamayo Goya
Esta especie muestra un crecimiento rápido, una longevidad corta y una talla de primera maduración pequeña. Tiempo medio de vida, corto, entre 2-3 años en el Mediterráneo y Atlántico andaluz y entre 4-5 años en el Atlántico gallego. El ejemplar más longevo observado se ha estimado en 6 años. La esperanza de vida aumenta en latitudes más altas, en aguas más frías. Corta longevidad si la comparamos con las de otros moluscos bivalvos, como por ejemplo (variaciones en función del autor del artículo y del hábitat de procedencia): berberecho, Cerastoderma edulis (Linnaeus, 1758), entre 7 y 14 años; chirla, Chamelea gallina (Linnaeus, 1758), entre 8 y 12 años; navaja, Ensis siliqua (Linnaeus, 1758), entre 10 y 12 años; mejillón, Mytilus edulis (Linnaeus, 1758), entre 15 y 24 años; ostra, Ostrea edulis (Linnaeus, 1758), entre 16 y 20 años; datil de mar, Litophaga litophaga, hasta 50 y 60 años. Los moluscos bivalvos más longevos datados son dos ejemplares de Arctica islandica (Linnaeus, 1767), 405 y 507 años respectivamente.
El análisis de las líneas, grietas, interrupciones o anillos de crecimiento externos e internos de la concha y las franjas hialinas (traslúcidas), permiten establecer la edad y estimar la tasa de crecimiento. Dado que no todas estas señas o huellas tienen el mismo significado, es necesario reconocer aquellas que tienen un carácter periódico, fenómenos estacionales, y los que tienen un carácter ocasional, como consecuencia de condiciones ambientales desfavorables, perturbaciones o estrés ambiental. (Imagen 2.12) Hay autores que consideran el crecimiento de esta especie como estacional, con dependencia básicamente de la variación de la temperatura del mar y la mayor o menor disponibilidad de alimentos.

Imagen 2.12. En estas instantáneas se aprecian mejor franjas hialinas, anillos, líneas y grietas de crecimiento; Las imágenes B/ y C/ corresponden a la A/ pero con un haz luminoso por la cara interior de la valva. Ejemplar de Valencia. España. Mar Mediterráneo. ©Jorge C. Tamayo Goya.
Las líneas de crecimiento anual son depositadas especialmente en otoño cuando la tasa de crecimiento se ralentiza coincidiendo con la disminución de la temperatura del agua de mar en la superficie. La frecuencia de franjas hialinas en el margen ventral se observa a finales de primavera y verano, coincidiendo con el período reproductivo de la especie, ciclo gametogénico, un proceso energético altamente exigente. La separación, más o menos espaciada, la frecuencia y la alternancia de las franjas o bandas hialinas muestran fenómenos o períodos estacionales. Alternancia de crecimiento activo un poco antes de la primavera y mitad del verano y más lento en el otoño y comienzo del invierno, períodos que dan como resultado franjas o bandas más traslúcidas o hialinas y opacas, respectivamente.
En algunas áreas de la concha las líneas de crecimiento externas son más apreciables mientras que en otras áreas son muy difíciles de distinguir. Observaciones han concluido que es posible que no haya formación de anillos o que no son apreciables en ejemplares de menos de 15 mm, mientras que las de más de 25 mm todos mostraron un anillo. Los anillos más recientes se depositan cerca unos de otros y cerca del margen ventral, lo que puede dificultar su distinción, especialmente si el margen está erosionado. Tienden a prolongarse a todo lo largo de la concha como resultado de una concentración progresiva y a menudo se asocian con una grieta en la concha. La distinción entre los distintos tipos de anillos resulta más evidente en individuos jóvenes que en los adultos, en los que el crecimiento más lento provoca que los anillos se encuentren muy cercanos entre sí o hasta que se sobrepongan. El falso anillo de crecimiento tiende a ser discontinuos y aunque también puede asociarse a una grieta en la superficie de la concha cuando se examina bajo un microscopio se caracteriza por la interrupción repentina del patrón de crecimiento normal. Las líneas internas son visibles en secciones delgadas cuando las conchas se cortan desde el umbo hasta el margen ventral.
La variable más utilizada por los autores que centran sus estudios en el crecimiento de esta especie es la longitud. Una longitud se asocia con un número determinado de anillos. Se puede efectuar una estimación mediante el método de conteo de bandas de crecimiento en cortes transversales de la concha que es copiado en “acetate peel”, que son réplicas de superficies grabadas incrustadas con ácido en una película, lámina, de acetato; para posteriormente ajustar al modelo de crecimiento de von Bertalanffy, que se utiliza ampliamente en biología marina para modelar la relación edad-talla de la mayoría de los peces y de los moluscos bivalvos. (Imagen 2.13)
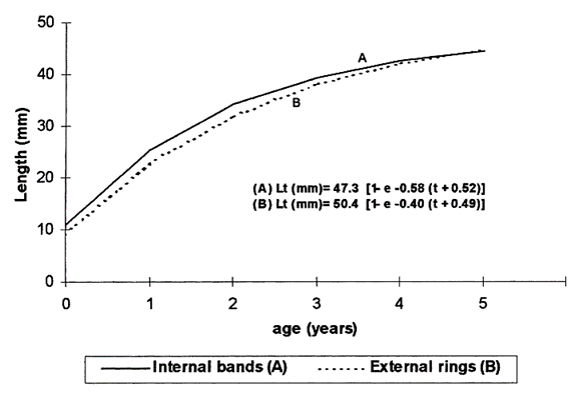
Imagen 2.13. Growth and reproductive cycle of Donax trunculus L., (Mollusca: Bivalvia) off Faro, southern Portugal. M.B. Gaspara, R. Ferreirab, C.C. Monteiroa. Una comparación de la tasa de crecimiento de D. trunculus determinada a partir de (A) réplicas de cáscara de acetato de secciones de caparazón y (B) superficie externa anillos de crecimiento. En esencia, este modelo parte de las siguientes observaciones: 1- El crecimiento no se produce a la misma velocidad a lo largo del tiempo, y 2- La velocidad de crecimiento inicial es muy rápida; después, a medida que aumenta de tamaño y madura sexualmente, la tasa de crecimiento va decreciendo poco a poco hasta ser prácticamente nula, deja de crecer. El estudio de estos incrementos regulares del crecimiento en las partes duras de los bivalvos y otros organismos calcificados se denomina esclerocronología.
• El cuerpo. Anatomía órganos internos.
El cuerpo, comprimido lateralmente, es de estructura simétrica bilateral, una parte de los órganos son dobles.
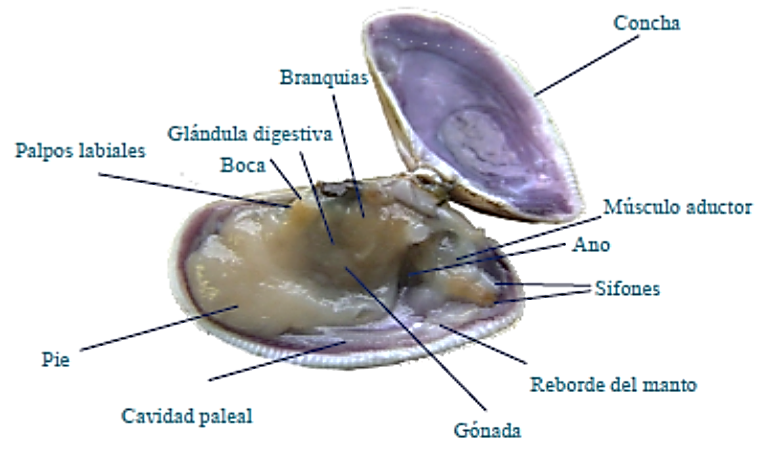
Imagen 2.14. Anatomía interna © Andrea Louzán.
El manto o palio, (Véase en la Imagen 2.22) es una formación tegumentaria, fina membrana carnosa, en el interior de ambas valvas. Es la que secreta la concha. Se compone de dos lóbulos simétricos, dos repliegues cutáneos, epitelio interno y externo, soldados uno al otro en el borde anterodorsal. Cada lóbulo está sujeto a su correspondiente valva por medio de los músculos paleales, insertos en la concha. El músculo orbicular del lóbulo interno deja una impresión, una visible línea paleal que sigue el contorno del margen de la concha. A la altura de los sifones, debajo del músculo aductor posterior, se produce un entrante en la línea paleal, llamado seno paleal. La cantidad y la posición de los músculos aductores, los que cierran las valvas, se utilizan también en la clasificación. Cuando hay dos, uno anterior y otro posterior, se denomina disposición dimiaria. El lóbulo interno del manto contiene el musculo orbicular que deja sobre la concha una impresión llamada línea paleal, también identificativa. Ambos son caracteres de la especie.
Es una proyección de la masa visceral que cubre las partes blandas del cuerpo. La masa visceral es la parte más voluminosa, tiene la forma de una bolsa en la que se alojan los órganos internos envueltos por el manto y, a su vez, protegidos por la concha: el aparato digestivo, sistema circulatorio, aparato excretor, sistema nervioso y órganos sensoriales, y aparato reproductor.
El espacio libre situado entre los dos lóbulos del manto y el cuerpo, la cavidad limitada por el manto, se denomina cavidad paleal o seno paleal (Véase en la Imagen 2.22). Por esta cavidad circulan las corrientes de agua que llevan el oxígeno, el alimento, arrastra los desechos y el anhídrido carbónico. El seno paleal es redondeado, profundo, un poco ascendente dorsalmente sobrepasando la mitad de la valva y confluente con la línea paleal, con el borde del manto.
En el borde del manto se aprecian tres pliegues (Imagen 2.6): el interno, desarrolla los músculos que le permite fijar el borde del manto a la concha; el intermedio, responsable de las funciones sensoriales; y el externo, que es el que secreta la concha. La glándula conchífera secreta inicialmente una única concha, se vuelve bivalva a medida que el manto se extiende.
La musculatura de los Bivalvos consta de tres sistemas principales, es decir, los músculos del manto o derivados de éste, los músculos de la concha y los músculos intrínsecos del pie y de la masa visceral.
Los bordes del manto están soldados salvo por tres aberturas, el pie (en el margen anterior), el sifón inhalante y el sifón exhalante (ambos en el margen posterior). (Imágenes 2.15, 2.16, 2.17, 2.19, 2.20 y 2.21) Los sifones, situados en la zona posterior, son prolongaciones del manto con forma de conducto tubular, de similar tamaño aunque difieren en el diámetro, grosor y en la cantidad y complejidad de los tentáculos sensoriales. Por el sifón inhalante, incurrente, ventral o branquial, situado más ventralmente, de mayor diámetro y con las paredes más gruesas, entra el agua portadora de oxígeno y alimento, materia orgánica en suspensión. Cuando el agua entra por el inhalante las papilas sensitivas, (Imagen 2.16) una corona de tentáculos pinnadamente ramificados dispuestos en tres series: 6 primarios, 6 secundarios y 12 terciarios, que rodean la abertura del sifón inhalante, realizan la primera filtración plegándolos hacia adentro formando un tamiz e impiden la entrada y expulsan en forma de pseudoheces las partículas demasiado grandes, como los granos de arena. Las heces y desechos de la digestión son expulsados, a la vez que sale el agua de la cavidad paleal, por la cámara y sifón exhalante, excurrente, dorsal o anal, que posee 12 tentáculos simples no ramificados y de menor diámetro que el sifón inhalante.
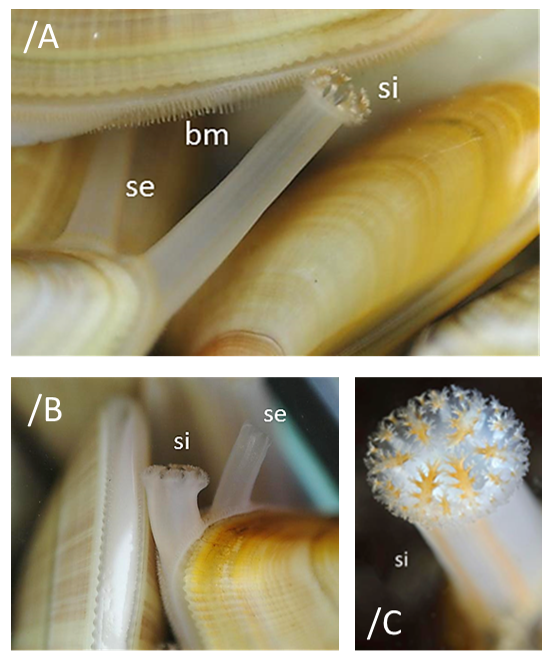
Imágenes 2.15/A y 2.16/B Detalle de sifones, borde del manto y tentáculos, órganos sensoriales. Borde del manto (bm); sifón exhalante (se); sifón inhalante (si). Ejemplares de Francia. Océano Atlántico. ©Cantabile. Imagen 2.17/C. Detalle del sifón inhalante (si) © Dugornay Olivier
La longitud de cada uno de los sifones se adecúa a las condiciones del medio, a sus necesidades fisiológicas y a la profundidad a la que se haya enterrado. La longitud del sifón podría determinar la profundidad máxima de enterramiento. En su máxima extensión pueden superar ampliamente en tamaño a la longitud de la propia concha. Cuando son amputados por el ataque de algún depredador pueden ser regenerados. Los músculos retractores son los que los proyectan o retraen y se alojan en la cavidad paleal.
En un estudio se señala que las especies del género Donax la alimentación se realiza más comúnmente abriendo el sifón de inhalación al ras de la superficie de la arena. (Imagen 2.18) Puede estar orientado hacia la costa o hacia las olas entrantes, probablemente dependiendo de las propiedades físicas del flujo de agua. El sifón de exhalación puede mirar en la misma dirección que el inhalante o puede mirar en la dirección opuesta. Entre ambos sifones extendidos hay separación para evitar que los desechos del exhalante puedan ser absorbidos por el inhalante.

2.18. Reconnaître les principaux bivalves fouisseurs ou foreurs au moyen de leurs siphons. Fig 102. © Yves MÜLLER.
El pie (Imágenes 2.19/A, 2.20/B y 2.21/C) es un órgano musculoso que se extiende fuera de las valvas. Grande, aproximadamente la mitad del tamaño corporal. Con forma de lengua puntiaguda en el extremo. Aplanado lateralmente, doblado al estar retraído. Es una prolongación de la masa visceral, cuelga de ella, una de las tres aberturas del manto (además de los dos sifones). Situado en la zona antero-ventral, cuelga en el interior de la cavidad paleal. Se proyecta desde el cuerpo hacia la cavidad del manto. Realiza una importante función excavadora que le permite hundirse en el fondo arenoso. En esta especie no desempeña función locomotora. Los músculos anterior y posterior lo fijan al interior de las valvas y le permiten una gran movilidad. La acción combinada de los músculos pedios, retractores y protractores, junto con la acumulación de la sangre, son los que lo extienden y lo retraen, a la vez que se contiene y se expulsa el agua de la cavidad paleal.
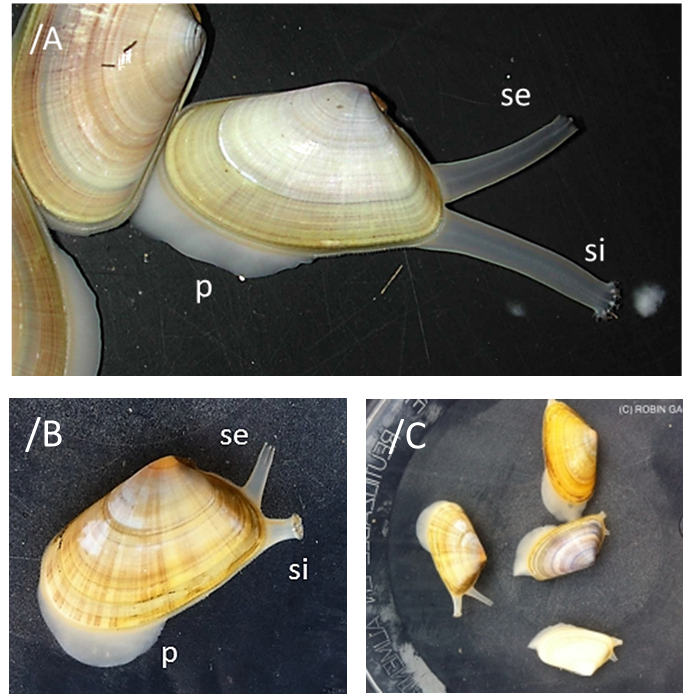
Detalle de ambos sifones y pie extendidos. Pie (p); sifón inhalante (si); sifón exhalante (se). Imagen 2.19/A Ejemplares de Huelva. España. Oceano Atlántico. © Junta de Andalucía. Imágenes 2.20/B y 2.21/C. Ejemplares de Sables d’Olonne. Francia. Océano Atlántico. © Robin Gauff.
Las dos voluminosas branquias laminares (ctenidios), lamelibranquias (eulamelibranquias), las más frecuentes y desarrolladas de los bivalvos, tienen forma de W, que con su continuo flujo de agua desempeñan dos funciones: respiratoria, toman oxígeno y desechan el dióxido de carbono, y nutricia, se nutren de alimentos. Cuelgan del techo de la cavidad paleal, en la cara interna del manto, a ambos lados de la masa visceral y a ambos lados del pie. (Imágenes 2.22 y 2.23) Dividen la cavidad del manto, cavidad paleal, en dos, una inferior o ventral (infrabranquial, corriente inhalante) y otra superior o dorsal (suprabranquial, corriente exhalante). Del eje superior de la branquia salen dos ramas eferentes y otras dos aferentes que la divididen en dos hemibranquias.
Filtran el alimento del agua, pueden incluso filtrar 30 litros al día. Lo dirigen hacia los palpos labiales en donde se efectúa la clasificación alimentaria antes de pasar a la boca. Las branquias retienen las partículas en suspensión, envolviéndolas en una capa de mucus y conduciéndolas hacia la boca. El bolo alimenticio, amalgamado por el moco, pasa a la boca o es rechazado por los palpos y eliminado por el animal en forma de las denominadas pseudoheces.
Imágenes 2.22 y 2.23
El aparato digestivo es una cavidad aplanada lateralmente de forma ovoide y globosa situado entre el esófago y el intestino. Las partículas alimenticias que previamente han pasado por los palpos labiales pasan por la boca (carente de mandíbulas) y por el corto esófago hasta el estómago. En el estómago, que se halla rodeado por el divertículo digestivo, se encuentra el estilo o estilete cristalino, una estructura en forma de varilla que tritura y disuelve las partículas mediante un movimiento giratorio y la producción de más mucus y de diversas enzimas digestivas que contribuyen a la digestión extracelular. La glándula digestiva o hepatopáncreas, órgano enlazado al estómago que hace las funciones del hígado y del páncreas, es en donde se efectúa la digestión intracelular. Se continúa por el intestino que presenta un complejo sistema de pliegues y es en donde se producen las heces, posteriormente por el recto después de atravesar el corazón y, finalmente, desemboca a través del ano en la parte posterior sobre el borde dorsal del músculo abductor, en la cámara exhalante para ser expulsadas por el sifón exhalante.
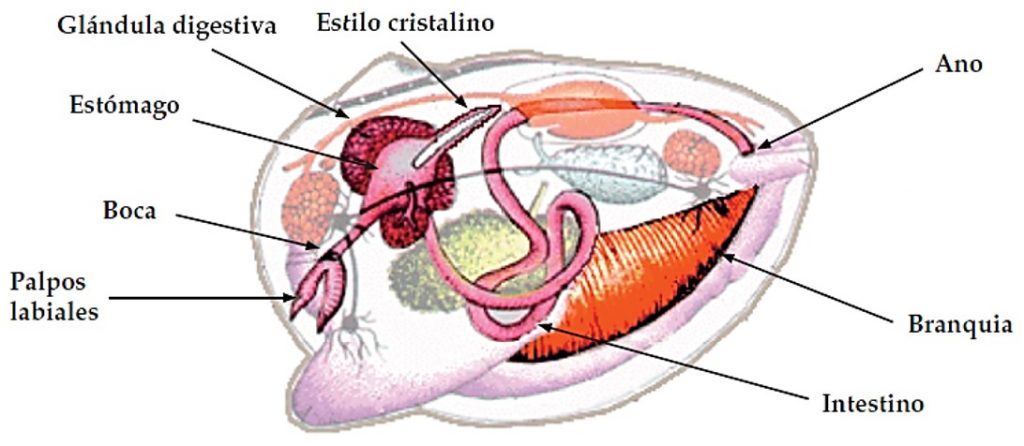
Imagen 2.24. Esquema del sistema digestivo de los bivalvos. ©CSIC
El sistema circulatorio o vascular sanguíneo, del corazón a los senos somáticos, a los órganos renales, a los respiratorios, y al corazón. Lo constituye un corazón envuelto por una fina membrana o saco transparente, pericardio, que rodea al intestino que a través el recto desemboca en el ano (como se ha señalado antes), fenómeno que se conoce como perforación del corazón. Posee dos aurículas laterales y un ventrículo. Por las aortas, una anterior y otra posterior, se distribuye la hemolinfa, que es un líquido incoagulable, incoloro, rico en una forma líquida de calcio que contiene hemocianina, que hace la función de la hemoglobina. Se distribuye a todas las partes del cuerpo a través de las arterias, arteriolas y los espacios libres en el tejido conjuntivo, denominados lagunas, circulando libremente antes de llegar al seno ventral. De aquí va al órgano excretor o riñón en donde se purifica y pasa a las branquias por medio de los vasos branquiales aferentes. Los dos pares de riñones o nefridios es en donde la hemolinfa es purificada para pasar luego a las branquias a través de los vasos branquiales aferentes. El par de riñones constituyen junto con las glándulas pericárdicas el aparato urinario, y otro par forman los gonoductos que finalmente se abren en el saco pericárdico para desembocar en la cavidad paleal.
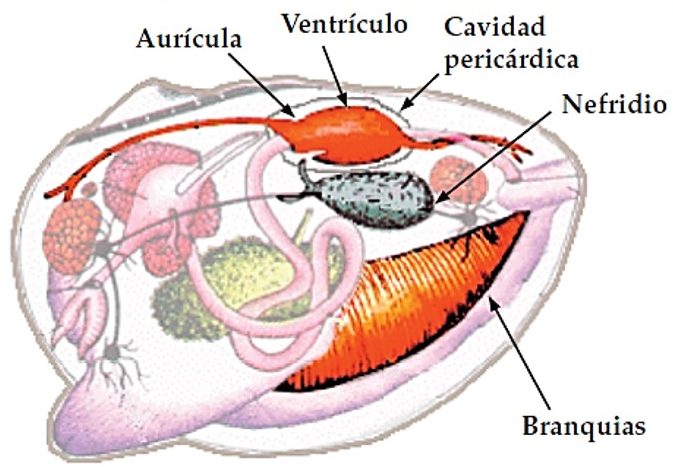
Imagen 2.25. Esquema del aparato circulatorio y excretor de los bivalvos. ©CSIC
El Sistema nervioso, bastante simple, simétrico bilateralmente, presenta tres pares de ganglios principales, son los cerebropleurales (actúan sobre el manto), viscerales (actúan sobre la masa visceral y las branquias) y pediales (actúan sobre el pie). Las regiones en donde se centraliza el mayor número de órganos sensoriales son los bordes del manto, ambos sifones y pie. Y están representados principalmente por estatoscitos, encargados del equilibrio, y osfradios, desarrollan funciones olfativas. En diversos puntos del cuerpo existen otros ganglios menos importantes, que son los previscerales.
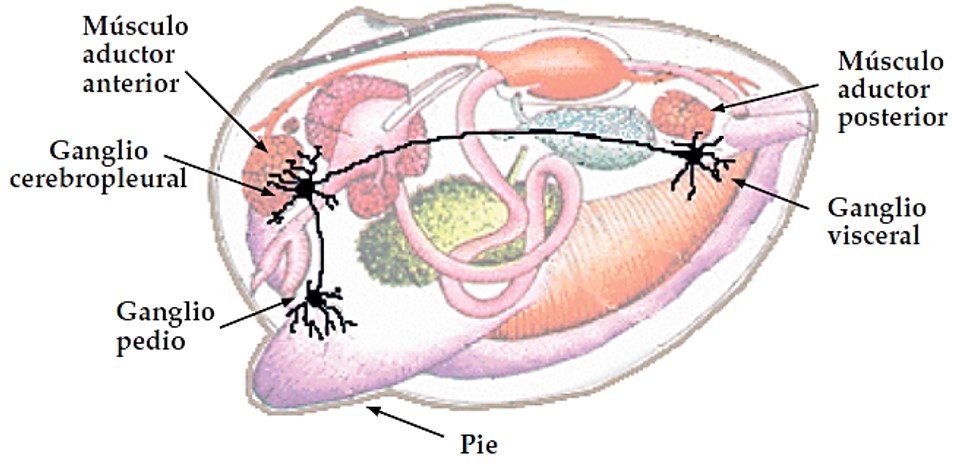
Imagen 2.26. Esquema del sistema nervioso, pie y músculos aductores de los bivalvos. ©CSIC
El aparato reproductor está formado por un par de gónadas y los conductos evacuadores. Se trata de una especie dioica, es unisexual, cada sexo se presenta en individuos diferentes y están representados en proporción bastante similar. No hay copulación. La producción de óvulos y esperma es un proceso denominado gametogénesis. La gónada, se encuentra en la masa visceral, es el órgano reproductor que emite gametos masculinos (espermatozoides) o femeninos (óvulos). Cuando la gónada comienza a desarrollarse el ovario adquiere una coloración azul oscuro en las hembras (rojiza en el caso de Donax vittatus y rosada en el caso de Donax semistriatus) mientras que el testículo del macho es blanquecino y anaranjado (Imágenes 2.27) En reposo sexual e individuo inmaduro, el sexo no es diferenciable, sexo indeterminado. Durante el período de actividad la gónada llega a ocupar gran parte de la masa visceral.
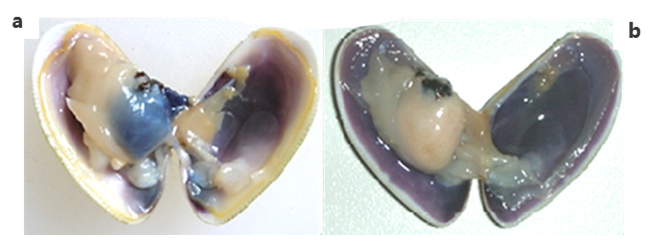
Imágenes 2.27. Gónadas período madurez. Hembra (a). Macho (b). ©Andrea Louzán Pérez.

La fecundación es externa. Expulsan los ovocitos y los espermatozoides al agua por el sifón exhalante. Los huevos fecundados al eclosionar generan larvas pelágicas (velígeras) que forman parte del plancton hasta que con el tiempo, ya desarrolladas, se establecen en el fondo. (Imágenes 2.28 y 2.29)
El desarrollo mediante fases larvarias tiene una vida pelágica en la que se alimenta de nanoplancton, entre tres o cuatro semanas. Al cabo de ese mes, aproximadamente, se produce la metamorfosis, desaparece el velo por completo, aparecen las branquias. En esta fase veliger es cuando aparece, cuando se aprecia mejor, la concha formada por dos valvas, secretadas por la glándula conchífera y posteriormente secretada por el manto, comienza a alimentarse exógenamente pasando de la vida pelágica (flotando en el mar) a la vida bentónica (en el fondo del mar). En la fase postlarva ya se aprecian pie, aparato digestivo y tejido branquial. El paso de larva a semilla supone el fin del desarrollo larvario. (Imágenes 2.28 y 2.29)
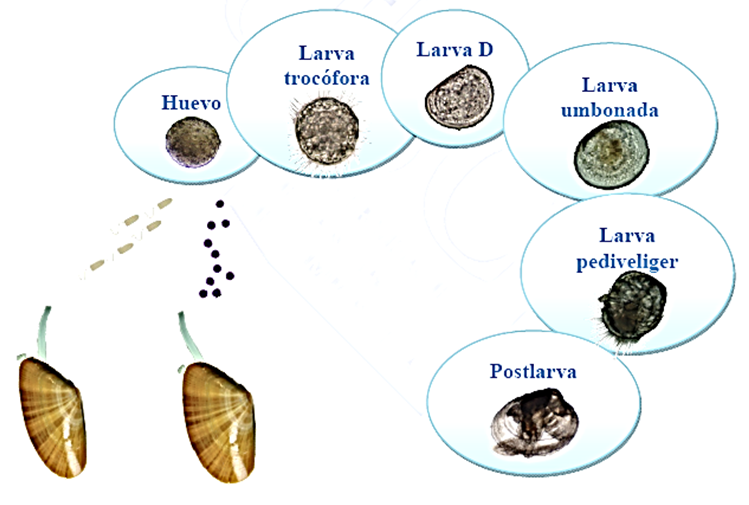
Imagen 2.28. Ciclo vital Donax trunculus. ©Andrea Louzán Pérez.
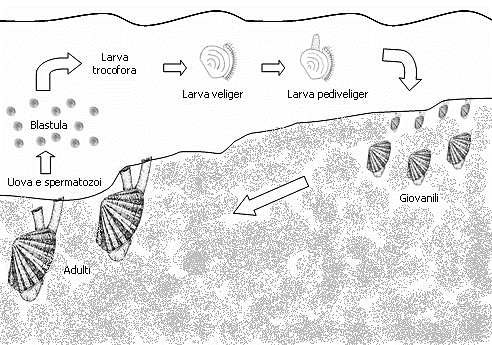
Imagen 2.29. Ciclo larvale di Donax trunculus. ©Paola La Valle
La reproducción en los bivalvos está sujeta a ciclos estacionales. Las épocas de madurez sexual y los ciclos reproductores están ligados a los factores medioambientales: temperatura, salinidad, pH y alimentación.
Aunque el desarrollo gonadal de Donax trunculus varía en función de la localización geográfica de los individuos, se puede indicar a modo de referencia que este comienza principalmente a partir de febrero y el desove a partir de la primavera con el aumento de la temperatura del agua (temperatura óptima en torno a 20º). Aunque es relativamente escalonada en el tiempo destacan dos fases principales: la primavera y el verano. La etapa más óptima es de marzo-abril a junio-julio. El desove estacional tiende a sincronizarse con condiciones ambientales óptimas que son fundamentales para la supervivencia y el aumento del reclutamiento. La fase de verano corresponde al final del período la actividad de la gónada que termina a final de otoño con la emisión masiva de gametos en junio-julio y septiembre-octubre. En estos períodos, además de los meses invernales, pueden llegar a perder entre el 17% y el 33% de su peso. Desde septiembre-octubre hasta diciembre-enero se produce una fase de reposo gonadal. Se apunta también otra fase en el verano con un agrupamiento completo entre Julio y agosto.
Se trata de una especie en la que trascurre un año entre nacimiento y madurez sexual, con una talla de primera maduración pequeña, amplio período de madurez y varios episodios de desove o hasta con abundantes puestas escalonadas en el tiempo, y un elevado número de huevos. A partir del año es posible la visualización macroscópica de la gónada desarrollada. Tanto machos como hembras muestran sincronismo en el desarrollo gonadal. Durante el período reproductivo muestra una liberación continua y simultánea de gametos y una alta capacidad de regeneración de los ovocitos. A menudo, hay una temporada de reproducción prácticamente ilimitada, aunque presentan picos de actividad reproductiva. Hay estudios que constatan que entre abril y julio esta especie mantiene al nivel más alto su capacidad de desove, 100% de los individuos. En una investigación, la fecundidad media realizada obtenidas de siete inducciones al desove en condiciones de laboratorio, se obtuvieron los siguientes valores de huevos puestos por hembra: 584.000, 812.00, 973.000, 948.000, 644.000, 1.030.000, 853.000.
La talla de la primera maduración sexual, cuando el 50% de los individuos están maduros, aunque varía en función de la autoría y de la zona costera en la que se haya realizado el estudio, es más frecuente establecerla entre 12 mm y 14 mm. Aunque también se señalan tallas menores en el Mediterráneo, de hasta 8 mm y 10 mm, y mayores en el Atlántico, de hasta 16 mm y 20 mm. La talla de maduración puede variar sensiblemente entre hembras y machos. Las clases de talla que acumulan el mayor porcentaje del potencial reproductivo son, en muchos casos, los de mayor tamaño, 28 mm y 30 mm, a partir de su segunda primavera-verano, entre 1,5 y 2 años de vida. La talla mínima de madurez sexual se puede establecer en 28 mm.
Todas las poblaciones naturales dependen en gran medida de su estructura y su dinámica, a partir de la contribución de nuevos individuos. Cuando esta contribución se debe a la reproducción sexual y no por migración, se define como reclutamiento. La estrategia de reclutamiento desarrollada por Donax trunculus muestra algunas diferencias en relación al área de distribución geográfica: bimodal, en dos períodos y unimodal, en un único período.
• Reseñas morfológicas, estudio de la forma y dimensiones. Alometría, relación entre el desarrollo de una parte del organismo y otra parte o con todo el conjunto. Los estudios muestran una mayor tendencia hacia la alometría negativa, crecimiento más lento, en la mayoría de las relaciones entre la longitud de la concha y los demás parámetros, en las relaciones largo/alto (L/H) y largo/ancho (L/Wd), aunque también hay estudios que muestran crecimiento alométrico positivo, crecimiento más rápido, en alguna etapa del desarrollo, como por ejemplo el exhibido en las relaciones peso gonadal/longitud de la concha y peso seco/longitud de la concha, que podría estar relacionado con muchos factores tales como disponibilidad de alimento, temperatura y madurez sexual.
